ماهیچه های اسکلتی یکی از سه نوع اصلی ماهیچه است ، دیگری ماهیچه قلب و ماهیچه صاف هستند.
بیشتر ماهیچههای اسکلتی توسط رشتههای کلاژن به نام تاندونها به استخوانها متصل میشوند.
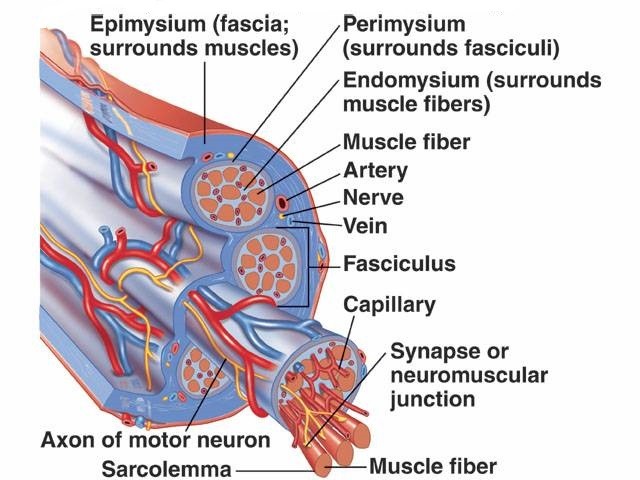
ماهیچه اسکلتی به دستههای متعددی از سلولهای متصل به هم به نام فیبرهای ماهیچهای تقسیم می شود.این الیاف و ماهیچهها توسط لایههای بافت به نام فاشیا احاطه شدهاند.
فیبرهای ماهیچهای و یا سلولهای ماهیچهای از ادغام میلوبلاست های رشته ای در فرآیندی به نام میوژنز تشکیل میشوند.
فیبرهای ماهیچهای استوانهای هستند و بیش از یک هسته دارند.
آنها همچنین چندین میتوکندری دارند تا نیازهای انرژی را برآورده کنند.
فیبرهای ماهیچهای به نوبه خود از میوفیبریل ها تشکیل شدهاند. میوفیبریل ها از رشتههای اکتین و میوسین تشکیل شدهاند که در واحدهایی به نام سارکومتر تکرار میشوند که واحدهای عملکردی اصلی فیبر عضلانی هستند.
سارکومر مسئول ماهیچههای اسکلتی است و اساس لازم برای انقباض ماهیچه را تشکیل میدهد.
فیبرهای ماهیچهای واحدهای انقباضی منحصر به فرد درون ماهیچه هستند. یک ماهیچه منفرد مثل عضله دو سر بازویی شامل فیبرهای ماهیچهای زیادی است.
گروه دیگری از سلولها، سلولهای میوساتللیت بین غشا پایه و فیبرهای ماهیچهای یافت میشوند. این سلولها به طور معمول ثابت هستند اما میتوانند با ورزش یا فیزیوتراپی فعال شوند تا میونوکلیی اضافی را برای رشد یا ترمیم ماهیچه فراهم کنند.
میکرو آناتومی
عضلات اسکلتی یک الگوی نواری مشخص را در زیر میکروسکوپ به خاطر آرایش عناصر اسکلت سلولی در سیتوپلاسم فیبرهای ماهیچهای نشان میدهد. پروتئینهای اصلی سیتوپلاسمی، میوزین و اکتین هستند (که به ترتیب به عنوان رشتههای “ضخیم” و “نازک” نیز شناخته میشوند) و در یک واحد تکرار شونده به نام سارکومر مرتب شدهاند. تعامل میوزین و اکتین مسئول انقباض ماهیچه است.
هر اندامک و ماکرومولکول فیبر ماهیچه برای اطمینان از این که فرم با عملکرد هماهنگ است، مرتب شدهاست.
غشا سلولی سارکوما نامیده میشود و سیتوپلاسم آن به سارکوپلاسم مشهور است. در میوفیبریل هاهستند.
میوفیبریل ها دستههای پروتئینی بلند با قطر حدود ۱ میکرومتر هستند که هر کدام حاوی میلوفیلامن هستند. با این که فیبرهای ماهیچهای فاقد شبکه اندو پلاسمیک نرم هستند اما حاوی شبکه اندو پلاسمیک گوشتی هستند.
شبکه سارکوپلاسمی میوفیبریل ها را احاطه کردهاست و ذخیره یونهای کلسیم مورد نیاز برای انقباض ماهیچه را در خود نگه میدارد. در فاصله بین دو دیواره انتهایی، یک تاخوردگی لولهای به نام لوله عرضی وجود دارد.

معماری ماهیچهای
معماری ماهیچهای به آرایش فیبرهای ماهیچهای نسبت به محور تولید نیرو در ماهیچه اشاره میکند. برای برخی ماهیچههای طولی مانند عضله دو سر بازویی، این مفهوم نسبتا سادهای است.
برای برخی دیگر مانند ماهیچه رکتوس فموریس یا دلتوئید، این امر پیچیدهتر میشود. با این که فیبرهای ماهیچهای یک فاسیکل به موازات یکدیگر قرار گرفتهاند. اما خود فاسیلها میتوانند در رابطه با یکدیگر و با تاندونهای خود متفاوت باشند. کششی که یک ماهیچه میتواند بین تاندونهای خود ایجاد کند، بیش از اندازه آن و آرایش فیبرها تغییر میکند.
معماری طولی
عضلات طولی مرتب شده، موازی و یا دوکی شکل موازی با محور تولید نیرو حرکت میکنند .در نتیجه این ماهیچهها در کل به طور مشابه با یک فیبر ماهیچهای بزرگ عمل میکنند. به عنون مثال یک ماهیچه دایرهای مانند اربیکولاریس است که در آن فیبرها به صورت طولی مرتب میشوند اما یک دایره از مبدا تا قرار دادن ایجاد میکنند.
معماری یکنواخت
فیبرها در ماهیچههای یکنواخت همگی در یک زاویه (اما غیر صفر) نسبت به محور تولید نیرو قرار میگیرند. این زاویه نیروی موثر هر فیبر را کاهش میدهد چون به طور موثر خارج از محور است.
با این حال، به خاطر این زاویه، فیبرهای بیشتری میتوانند در حجم ماهیچه یکسان بستهبندی شوند و سطح مقطع فیزیولوژیکی (PCSA را افزایش دهند. به طور کلی سرعت کوتاه شدن ماهیچه در مقایسه با سرعت کوتاه شدن فیبر همانند کل فاصله کوتاه شدن کاهش مییابد.
همه این اثرات با زاویه پنائوس مقیاس میشوند. واستوس لترالیس نمونهای از معماری یکپارچه یا یکنواخت است.
معماریهای چند نقطهای
فیبرها در ماهیچههای چند قسمتی در زوایای چندگانه نسبت به محور تولید نیرو مرتب میشوند ودر واقع عمومیترین و رایجترین معماری هستند. چندین جهت گیری فیبر در این گروه قرار میگیرند.
تغییرات گسترده زوایای پنتان در این معماری در واقع میتواند اجازه توابع چندگانه را بدهد. به عنوان مثال، عضله ذوزنقه، یک ماهیچه همگرا اولیه، میتواند هم در بالا بردن شانه و هم در افسردگی کمک کند.
ترتیبات چند قسمتی به یک آرایش خاص محدود نمیشوند .بلکه هنگامی که به طور خاص مورد استفاده قرار میگیرند به چیزی اشاره میکنند که اساسا ترکیبی از ترتیبات دو قسمتی یا تک قسمتی با ترتیبات همگرا است.
فیزیولوژی سلولی و انقباض
علاوه بر اجزای اکتین و میوزین که سارکومر را تشکیل میدهند، فیبرهای عضلانی اسکلتی نیز شامل دو پروتئین تنظیمی مهم دیگر یعنی تروپونین و ترومیوزین هستند که برای انقباض ماهیچه ضروری هستند.
این پروتیینها با اکتین مرتبط هستند. و برای جلوگیری از تعامل آن با میوزین همکاری میکنند. سلولهای ماهیچه اسکلت قابل تحریک هستند و توسط استیل کولین انتقالدهنده عصبی که در محل اتصال عصب – عضله توسط نورونهای حرکتی آزاد میشود، دپولاریزاسیون میشوند.
وقتی یک سلول به اندازه کافی تحریک میشود، شبکه سارکوپلاسمی سلول، کلسیم یونی (Ca۲ +)آزاد میکند و سپس با پروتئین تنظیمی تروپونین تعامل میکند. تروپونین متصل به کلسیم تحت یک تغییر ساختاری قرار میگیرد که منجر به حرکت تروپیمیوزین میشود و متعاقبا جایگاههای اتصال میوزین به اکتین آشکار میشوند. این کار چرخه پل عرضی وابسته به مایکوزین و اکتین ATP و کوتاه شدن ماهیچه را ممکن میسازد.
فیزیک
نیروی ماهیچهای متناسب با سطح مقطع فیزیولوژی (PCSA) است .و سرعت ماهیچه متناسب با طول فیبر ماهیچهای است.
با این حال، اطراف یک مفصل با تعدادی از پارامترهای بیومکانیکی، از جمله فاصله بین قسمتهای داخلی ماهیچه و نقاط محوری، اندازه عضله و نسبت دنده معماری تعیین میشود.
ماهیچهها به طور نرمال در مقابل هم چیده میشوند به طوری که وقتی یک گروه از ماهیچهها منقبض میشوند، گروه دیگر استراحت میکند.
در طول حرکات بالستیکی مانند پرتاب، ماهیچههای مخالف در طول انقباض، به ویژه در پایان حرکت، به ماهیچههای اصلی ترمز میکنند. در مثال پرتاب، سینه و جلوی شانه (دلتوئید قدامی)منقبض میشوند تا بازو را به سمت جلو بکشند، در حالی که ماهیچههای پشت و پشت شانه (دلتوئید خلفی)نیز منقبض میشوند و تحت انقباض برونگرا قرار میگیرند تا حرکت را آهسته کنند تا از آسیب جلوگیری شود.
بخشی از فرآیند تمرین، یادگیری ریلکس کردن ماهیچههای مخالف برای افزایش نیروی ورودی سینه و شانه جلویی است.
انقباض ماهیچهها تولید ارتعاش و صدا میکنند. فیبرهای کششی کند ۱۰ تا ۳۰ انقباض در ثانیه ایجاد میکنند. فیبرهای انقباض سریع ۳۰ تا ۷۰ انقباض در ثانیه ایجاد میکنند.


